
Résumé – Le status des Cytilus Européens est présenté puis discuté à la lumière de nouvelles données d’écologie de morphologie et de l’analyse de séquences d’ADN mitochondrial. La synonymie entre Cytilus sericeus (Forster, 1771) et C. auricomus (Duftschmid, 1825) ne semble pas justifiée ce qui conduit à proposer la réhabilitation de C. auricomus comme bonne espèce [stat. nov.]. Une clé d’identification est donnée pour distinguer les deux Cytilus d’Europe.
There are indeed two species of Cytilus in Europe
Abstract – The status of European Cytilus is presented and discussed in the light of new ecological and morphological data and the analysis of mitochondrial DNA sequences. The synonymy between Cytilus sericeus (Forster, 1771) and C. auricomus (Duftschmid, 1825) does not appear justified, which leads to propose the rehabilitation of C. auricomus as a good species [stat. nov.]. An identification key is given to distinguish the two European Cytilus.
Contexte
Au sein des Byrrhidae (Coleoptera) à élytres striés (Byrrinae), le genre Cytilus, se distingue des Byrrhus par les tibias moyens et postérieurs incomplètement sillonnés (le tarse ne se replie pas en entier dans le sillon), tandis que ces mêmes pattes ne disposent pas de fossettes où se loger au repos (Lompe 2015).
Pour ce genre, c’est d’abord l’espèce Byrrhus varius de Fabricius (1775: 60) qui sera utilisée avant d’être considérée à juste titre comme synonyme junior du C. sericeus de Forster (1771: 16). Ce dernier décrit Cistela sericea à élytres vert-brillants et interstries interrompues par des taches et soies noires. Duftschmid (1825 p. 16-17), séparera Byrrhus auricomus par la couleur du dessus foncièrement sombre, sans reflets verts, et avec une couverture homogène de soies gris-dorées.
Néanmoins, la coloration peut être problématique pour identifier les Byrrhidae, dans la mesure où les individus sont souvent frottés et leurs soies absentes (FIG. 4 à gauche), ou encore qu’il peut exister une certaine variation. Ainsi, pour C. sericeus, de nombreux types de coloration et de vestiture sont détaillés par Erichson (1845-1848 p. 490-491), qui à la suite les considère tous comme variétés, sans apporter de caractères complémentaires. Tout au plus, la forme plus étroite du C. auricomus est indiquée.
Czwalina (1878) est le premier à détailler l’aspect des sternites. Chez les Cytilus d’Europe, chaque sternite est en effet bordé sur toute sa largeur d’une micro-réticulation très serrée, d’aspect mat, plus ou moins large (FIG. 5 et FIG. 6). Czwalina indique que cette bande mate couvre la seule moitié postérieure de chaque sternite chez C. sericeus, la totalité de chaque sternite chez C. auricomus. Pour cet auteur, ce caractère est constant et les éléments associés également (colorations, étroitesse du corps). Il rapporte plusieurs individus découverts dans les zones moussues des marais liés au fleuve Pregel près de Königsberg (Prusse), lieu qui correspond actuellement au fleuve Pregolia à Kaliningrad (Russie). Il considère la « forte pilosité abdominale » [= microréticulation] comme une adaptation à une forte humidité.
Pour ses clés d’identifications, Reitter (1881 p. 80) va reprendre les informations de Czwalina mais le caractère relatif à l’aspect des sternites va disparaître sans justification dans mise à jour suivante (Reitter 1911 p. 160).
Letzner (1884) apporte de la confusion en confirmant les caractères des sternites mais, en se focalisant sur les variantes de coloration et de pilosité, il ne fera qu’allonger la liste des variétés établie par Erichson tout en émettant des doutes sur la valeur spécifique de C. auricomus.
Dans sa faune du Danemark, Hansen (1938) sépare les deux espèces en intégrant les éléments des sternites, puis à sa suite H.F. Paulus (in Freude et al. 1979). Ces deux clés d’identifications ont servi de référence jusqu’à ce que A. Pütz (in Lucht & Klausnitzer 1998) ne considère, sans justification évidente, que C. auricomus est synonyme de C. sericeus : « auricomus ist eine Variation von sericeus (Forster, 1771) und als Synonym dieser Art anzufügen« . Cette action va ainsi retirer l’espèce des inventaires subséquents (O. Jäger & A. Pütz in Löbl & Smetana 2006, in Löbl & Löbl 2016).
Il est à présent utile de rappeler la note de R. Allemand « Le taxon auricomus Duftschmidt, 1825 : 16 (Byrrhus) a été placé en synonymie sans justification alors qu’il paraît être une espèce distincte à la fois sur des critères morphologiques (fugaces), moléculaires (Allemand et al., non publié) et biologiques (tourbières, marécages) » (R. Allemand in Tronquet & Peslier 2024). La publication évoquée ici n’a pas connu de suite en dehors d’un référencement dans GenBank.
Habitats
Lors de mes inventaires dans les Alpes, j’ai pu collecter de nombreux Cytilus dont une partie correspond clairement aux descriptions du C. auricomus. Dans mes données, C. sericeus est largement distribué et se rencontre dans divers habitats moussus, qu’ils soient sur des sols humides, en bord de ruisseaux, y compris en altitude, ou encore au pieds des arbres. En revanche, C. auricomus ne m’est connu que de tourbières et de cœur de marais en zone facilement inondable, ce qui constitue un habitat très particulier et contraignant. À mon sens, cela est particulièrement cohérent avec les indications de forte rareté et d’une distribution limitée en Europe Centrale (Duftschmid 1825, Czwalina 1878, Reitter 1911).
ADN mitochondrial
Les bases de données d’ADN mitochondrial de coléoptères se sont mises en place récemment (Pentinsaari et al. 2014, Hendrich et al. 2015, Rougerie et al. 2015). Elles peuvent être particulièrement utiles pour les cas comme celui qui nous intéresse (Vuataz et al. 2019, Germann et al. 2020, Raupach et al. 2020).
La base de données BOLD (Ratnasingham & Hebert 2007, https://v3.boldsystems.org/) regroupe en 2024 pour les Cytilus sericeus et auricomus, 22 séquences exploitables de la Cytochrome Oxidase sous-unité 1 région 5′. La base GenBank (www.ncbi.nlm.nih.gov) en ajoute 3 de plus.
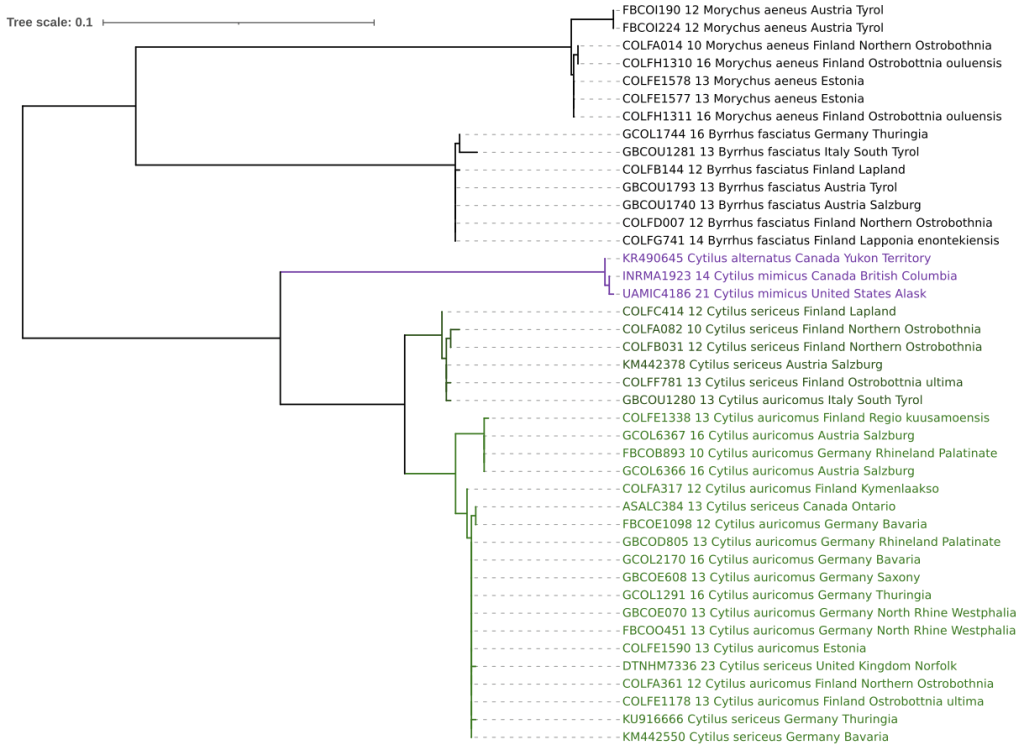
J’ai utilisé comme groupes extérieurs deux espèces largement distribuées et proches taxonomiquement : Byrrhus fasciatus (Forster, 1771) et Morychus aeneus (Fabricius, 1775). Parmi les séquences disponibles pour le genre Cytilus, figurent deux taxons du nord américain : Cytilus alternatus (Say, 1825) et C. mimicus Casey, 1912, qui ont été sélectionnés ici. L’analyse des séquences est automatisée (Lemoine et al. 2019, https://ngphylogeny.fr/, Letunic & Bork 2024, https://itol.embl.de).
Les divergences entre le groupe Cytilus sericeus – auricomus et les groupes externes sont importantes : supérieures à 21,40 % si l’on considère Byrrhus fasciatus, et à plus de 23,62 % par rapport à Morychus aeneus.
Entre les deux Cytilus du groupe sericeus – auricomus, les divergences relevées s’échelonnent de 0,00 % à 4,49 %, avec deux groupes bien délimités (FIG. 1).
Comme attendu, le duo Cytilus alternatus – mimicus se rattache au genre Cytilus mais avec une position distincte des deux autres espèces. Une synonymie entre les deux duos est ainsi exclue. En revanche, les deux taxons américains sont mal différentiés et selon les auteurs, regroupés sous C. alternatus ou scindés en espèces distinctes (cf. Johnson 1991). L’absence de divergence entre eux obtenue ici (0,35 %) viendrait valider un statut unique mais avant cela, il faudrait en savoir plus sur les spécimens et ces espèces.
Un autre Cytilus canadien (Ontario), identifié comme C. sericeus est regroupé avec les « C. auricomus » d’Europe (spécimen ASALC384, 2e groupe de la FIG. 1). La présence de cette espèce en Amérique du nord a déjà été indiquée (Takizawa & Nakane 1977, Tshernyshev 2006), mais pas celle de C. auricomus. D’autre part, deux spécimens ont pu être vérifiés grâce aux photographies des sternites, l’un est correctement nommé C. sericeus (COLFA082-10, Finlande, 1er groupe de la FIG. 1), le second, nommé C. sericeus (DTNHM7336-23, Royaume-Uni, 2e groupe de la FIG. 1), correspond en fait à C. auricomus et semble ainsi correctement placé dans l’analyse.
Les autres spécimens incohérents du groupe Cytilus sericeus – auricomus seraient donc à revoir directement pour assurer leur identification, car les critères utilisés ou le concept des sous-espèces retenu n’ont pas été précisés, tandis que les photographies ne montrent qu’exceptionnellement les sternites. Il s’agit d’un possible C. sericeus (GBCOU1280-13, Tyrol Italien), et de deux possibles C. auricomus (KU916666 et KM442550, Allemagne).
Morphologie
Mes observations ont été faites sur 11 spécimens de C. auricomus et 69 de C. sericeus, dont particulièrement 3 de chaque type : C. auricomus de la tourbière du Lac Luitel, Séchilienne, 38 (à gauche sur les images ci-après), et C. sericeus de la Montagne du Saut, Les Allues, 73 (à droite).
La coloration verte métallique est évidente pour la plupart des C. sericeus mais s’avère indistinguable chez certains spécimens, ce qui vient ainsi les rapprocher de l’aspect des C. auricomus (FIG. 2 à droite). En revanche, les interstries paires présentent bien, chez C. sericeus, les groupes de soies noires structurées en damier tel que rapporté dans la littérature. Chez C. auricomus la pilosité est homogène, et lorsque des soies noires sont présentes, elles ne forment pas de taches denses en damier.
La largeur de l’élytre est légèrement moindre chez C. auricomus mais cela s’apprécie mieux par comparaisons de photographies prises sous le même angle et avec des mesures par informatique. Sous la loupe, la différence n’est pas évidente à déceler.

La largeur relative de l’élytre est indiquée
Il faut aussi noter que les élytres sont légèrement écartées, ce qui impacte la forme générale
Le second critère observé concerne la ponctuation. Au niveau de la tête, elle est légèrement plus forte chez C. sericeus mais, à nouveau ce caractère n’est pas évident sans comparaison directe (FIG. 3).

La ponctuation du pronotum offre le même résultat, avec des points un peu plus forts chez C. sericeus. La densité semble identique (FIG. 4). Ce critère ne me semble pas fiable pour aider à l’identification. Au niveau des élytres aucune différence notable de ponctuation n’a pu être découverte.

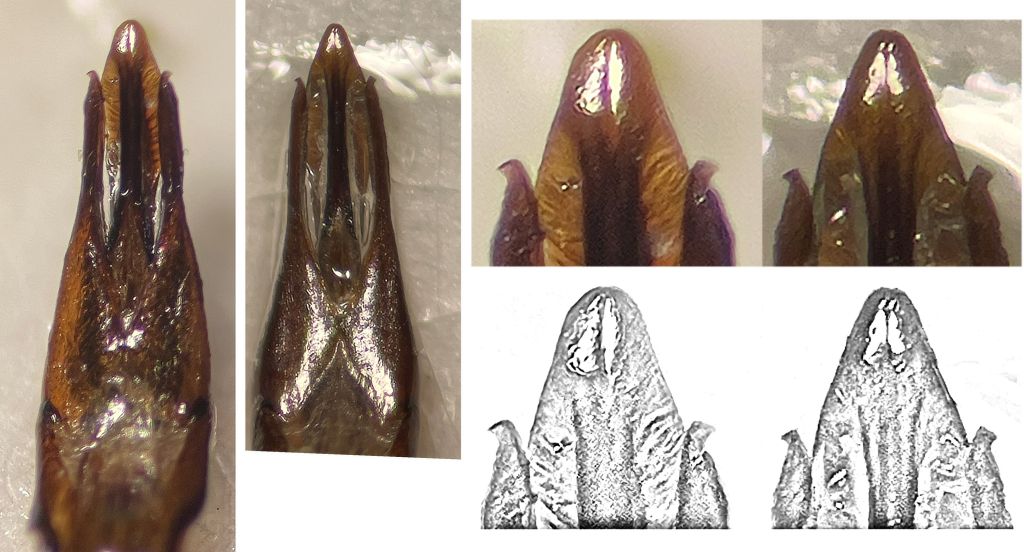
La microréticulation des sternites est le caractère discriminant le plus évident entre les deux taxons. Les espaces brillants et mats sont aisés à différentier (FIG. 5, FIG. 6). Je n’ai pas constaté de dispositions intermédiaires ni de variantes.
L’aspect mat n’est pas lié à de micro-soies appliquées comme cela a pu être indiqué. Il s’agit bien d’une microréticulation iso-diamétrale bien gravée mais très serrée, visible à grossissement x70.

La retouche en noir et blanc montre plus nettement les zones mates et brillantes chez C. sericeus (droite) et sur le premier sternite de C. auricomus (gauche)

Le dernier élément observé est l’édéage, dont les formes et proportions générales sont particulièrement semblables entre les deux taxons. Ce n’est qu’à l’examen côte-à-côte des clichés agrandis que l’on peut percevoir un apex du pénis plus large et plus arrondi chez C. auricomus tandis que l’apex des paramères se termine en crochet plus étroit et plus pointu chez C. sericeus. Ces caractères restent donc subtiles, tout comme en vue de profil où aucune différence n’a été détectée.
Discussion
Habituellement, la divergence entre espèces est placée au-dessus de 2,2 % (Ratnasingham & Hebert 2013, Rougerie et al. 2015). La valeur de 4,49 % obtenue ici laisse donc une large place à l’existence de deux espèces qui viennent corroborer la partition en deux groupes.
Des éléments autres que la présence d’espèces cryptiques peuvent intervenir sur le taux de divergence et en premier lieu, des problèmes liés à l’échantillonnage (Mutanen et al. 2016). Viennent ensuite, avec des proportions de l’ordre de l’exceptionnel, la présence d’ADN d’organismes symbiotes comme les Wolbachia (Smith et al. 2016), ou encore les copies de gènes surnuméraires (Raupach et al. 2016). N’ayant pas prise sur les échantillonnages, il m’est impossible d’enquêter plus avant pour établir leurs impacts éventuels sur les données traitées ici et l’on ne peut que s’en remettre à l’expertise des auteurs sources pour les aspects d’échantillonnage.
Nous avons donc au sein des Cytilus du groupe sericeus – auricomus :
- Deux groupes distincts, offrant des divergences génétiques suffisantes pour valider deux espèces ;
- Le caractère de la microréticulation des sternites qui est évident et stable pour plusieurs auteurs et parmi nos spécimens ;
- La présence ou l’absence bien tranchée de groupes de soies noires sur les élytres ;
- Des différences de ponctuation subtiles, difficiles à interpréter sans spécimens de référence ni photographies ;
- Des différences au niveau de l’édéage tout aussi subtiles ;
- La présence de C. auricomus dans les tourbières et marais avec des populations sporadiques en Europe Centrale alors que C. sericeus est largement distribué dans divers habitats moussus ;
- Des clés d’identifications historiques principalement basées sur la coloration dont la forte variabilité est connue pour C. sericeus ce qui ne facilite pas la détection de C. auricomus.
Ces éléments m’incitent à proposer la réhabilitation de Cytilus auricomus (Duftschmid, 1825) comme bonne espèce (stat. nov.).
Clé d’identification
- Sternites 2-4 à partie médiane lisse et brillante, la partie apicale étant mate du fait d’une microréticulation très fine et très dense. Sternite 5 entièrement brillant. Élytres à reflets métalliques verts présents ou atténués ; présence de taches de soies noires disposées en damier. Édéage plus étroit à l’apex avec les paramères étroits et pointus au sommet. Mousses d’habitats très variés. Large distribution, assez commun.
……….Cytilus sericeus (Forster, 1771). - Sternites 2-4 ainsi que le 5e entièrement mats du fait d’une microréticulation isodiamétrique très fine et très dense. Élytres plus ternes, à faible reflet métallique ; soies gris-dorées sans larges taches de soies noires. Édéage plus large à l’apex avec les paramères larges et arrondis au sommet. Semble exclusif des tourbières. Large distribution mais sporadique.
……….Cytilus auricomus (Duftschmid, 1825).
Données vérifiées pour C. auricomus
- Séchilienne (38), Lac Luitel, pièges Barber Prb et MB, 2 spécimens le 26/05/2011 ; piège Barber 4B, 6 spécimens entre le 02/06 et le 02/07/2014 ; piège Barber 6B, 1 spécimen le 02/07/2014 ; coll. B. Dodelin ;
- Saint-Jean-de-Chevelu (73), Marais de Chevelu, piège Barber PH, 1 spécimen le 09/06/2017 ; coll. B. Dodelin ;
- Thorens-Glières (74), Tourbière de Champ Laitier, piège Barber, 1 spécimen le 30/05/2023 ; coll. B. Dodelin.
Les données vérifiées de C. sericeus dont je dispose portent sur 69 spécimens distribués dans les départements 05, 38, 73, 74, 76 et 77.
Un nouveau Morychus ?
En marge des analyses spéciales aux Cytilus, les divergences intraspécifiques relevées ici sont au maximum de 0,82 % pour pour Byrrhus fasciatus ce qui n’appelle pas de commentaires particuliers, mais en revanche, de 2,08 % pour Morychus aeneus. Deux spécimens provenant du Tyrol Autrichien sont porteurs de cette divergence. Les Morychus de cette région, et peut-être plus largement des Alpes, seront donc à étudier en détail car ils pourraient constituer une espèce ou sous-espèce inédite.
Autorisations
Plusieurs spécimens indiqués ici proviennent d’espaces naturels protégées (Lac Luitel, Montagne du Saut dans le Parc national de la Vanoise). Toutes les captures ont été effectuées avec autorisation et mandat du gestionnaire.
Bibliographie
Allemand, R., Henri, H., Lemaitre, C. & Fabbri, R. (2010) Phylogenetic analysis of some Palaearctic Byrrhids (Coleoptera: Byrrhidae). Unpublished.
Czwalina, G. (1878) Cytilus auricomus Dft. Deutsche entomologische Zeitschrift, 22 (1): 205-206.
Duftschmid, C.E. (1825) Fauna Austriae. Oder Beschreibung der österreichischen Insecten, für angehende Freunde der Entomologie. – Linz, Akad. Buchhandlung – Bd. 3 [2]+1-289 https://doi.org/10.5962/bhl.title.51785
Erichson, W.F. (1845-1848) Naturgeschichte der Insecten Deutschlands. Erste Abtheilung. Coleoptera. Bd.3 Abt.1: Scaphidiidae – Scarabaeidae. Berlin, Nicolai’sche Buchhandlung, 968 pp. www.biodiversitylibrary.org/item/35217
Fabricius, J.C. (1775) Systema Entomologiae sistens Insectorum classes, ordines, genera, species, adjectis synonymis, locis, descriptionibus, observationibus. Flensburgi et Lipsiae, Korte [30]+832 p.
Forster, J.R. (1771) Novae species Insectorum. Centuria I. – Londini, T. Davies et B. White VIII+100 p.
Freude, H., Harde, K.W., & Lohse, G.A. (1979) Kafer Mitteleuropas. Bd 6. Diversicornia (Lycidae – Byrrhidae). Spektrum Akademischer Verlag, 367 pp.
Germann, C., Wyler, S. & Bernasconi, M.V. (2020) DNA barcoding of selected alpine beetles with focus on Curculionoidea (Coleoptera). Revue suisse de Zoologie, 124 (1): 15-38. https://doi.org/10.5281/zenodo.322661
Hansen, V. (1938) Danmarks fauna. Illustrerede haandbøger over den danske dyreverden. Bd.44 [Biller X Blødvinger, Klannere]. Dansk naturhistorisk forening. 320 pp. www.biodiversitylibrary.org/item/111380
Hendrich, L., Morinière, J., Haszprunar, G., Hebert, P.D.N., Hausmann, A., Köhler, F. & Balke, M. (2015) A comprehensive DNA barcode database for Central European beetles with a focus on Germany: adding more than 3500 identified species to BOLD. Mol Ecol Resour, 15: 795-818. https://doi.org/10.1111/1755-0998.12354
Johnson, P.J. (1991) Taxonomic notes, new records, and a key to the adults of North American Byrrhidae (Coleoptera). Proceedings of the Entomological Society of Washington, 93 (2): 322-332.
Lemoine, F., Correia, D., Lefort, V., Doppelt-Azeroual, O., Mareuil, F., Cohen-Boulakia, S. & Gascuel, O. (2019) NGPhylogeny.fr: new generation phylogenetic services for non-specialists. Nucleic Acids Research, 47: W260–W265. doi: 10.1093/nar/gkz303
Letunic, I. & Bork, P. (2024) Interactive Tree of Life (iTOL) v6: recent updates to the phylogenetic tree display and annotation tool. Nucleic Acids Research, gkae268 : 1-5. https://doi.org/10.1093/nar/gkae268
Letzner, K. (1884) Über Cistela (Cytilus) varia F. und auricoma Duft. Jahres-Bericht der Schlesische Gesellschaft für Vaterlandische Kultur, 61 [1883]: 302-304. www.biodiversitylibrary.org/item/176988
Löbl, I. & Löbl, D. (Eds) (2016) Catalogue of Palaearctic Coleoptera. Volume 3. Revised and updated edition. Scarabaeoidea – Scirtoidea – Dascilloidea – Buprestoidea – Byrrhoidea. Brill, Boston, 983 pp.
Löbl, I. & Smetana, A. (Eds) (2006) Catalogue of Palaearctic Coleoptera. Volume 3. Scarabaeoidea – Scirtoidea – Dascilloidea – Buprestoidea – Byrrhoidea. Apollo Books. Stenstrup, 690 pp.
Lompe, A. (2015) Käfer Europas – Familie Byrrhidae. https://coleonet.de/coleo/texte/byrrhidae.htm
Lucht, W. & Klausnitzer, B. (1998) Die Käfer mitteleuropas. Band 15: Supplementband 4. Goecke & Evers, Krefeld, 398 pp.
Mutanen, M., Kivelä, S.M., Vos, R.A., Doorenweerd, C., Ratnasingham, S., Hausmann, A., Huemer, P., Dincă, V., van Nieukerken, E.J., Lopez-Vaamonde, C., Vila, R., Aarvik, L., Decaëns, T., Efetov, K.A., Hebert, P.D.N., Johnsen, A., Karsholt, O., Pentinsaari, M., Rougerie, R., Segerer, A., Tarmann, G., Zahiri, R. & Godfray, H.C.J. (2016) Species-Level Para- and Polyphyly in DNA Barcode Gene Trees: Strong Operational Bias in European Lepidoptera, Systematic Biology, 65 (6): 1024–1040. https://doi.org/10.1093/sysbio/syw044
Pentinsaari, M., Hebert, P.D.N. & Mutanen, M. (2014) Barcoding Beetles: A Regional Survey of 1872 Species Reveals High Identification Success and Unusually Deep Interspecific Divergences. PLoS ONE, 9 (9): e108651. doi:10.1371/journal.pone.0108651
Pütz, A. (1998) Familie Byrrhidae. pp. 238-239 in Lucht, W. & Klausnitzer, B. (éd.): Die Käfer Mitteleuropas, 15.
Ratnasingham, S. & Hebert, P.D.N. (2007) BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Molecular Ecology Notes, 7: 355-364. https://doi.org/10.1111/j.1471-8286.2007.01678.x
Raupach, M.J., Hannig, K., Morinière, J. & Hendrich, L. (2016) A DNA barcode library for ground beetles (Insecta, Coleoptera, Carabidae) of Germany: The genus Bembidion Latreille, 1802 and allied taxa. ZooKeys 592: 121-141. https://doi.org/10.3897/zookeys.592.8316
Raupach, M. J., Hannig, K., Morinière, J., & Hendrich, L. (2020) A DNA barcode library for ground beetles of Germany: the genus Agonum Bonelli, 1810 (Insecta, Coleoptera, Carabidae). Deutsche Entomologische Zeitschrift, 67 (2): 197-207.
Reitter, E. (1881) Bestimmungs-Tabellen der europäischen Coleopteren. 4. Heft. Cistelidae, Georyssidae und Thorictidae. Verhandlungen der Kaiserlich-Königlichen Zoologisch-Botanischen Gesellschaft in Wien, 31: 67—96, 1 Taf. http://www.biodiversitylibrary.org/item/48128
Reitter, E. (1911) Fauna germanica: Die Käfer des Deutschen Reiches. Bd. 3. Schriften des Deutschen Lehrervereins für Naturkunde. 436 pp. & pl. 81-128.
Rougerie, R., Lopez-Vaamonde, C., Barnouin, T., Delnatte, J., Moulin, N., Noblecourt, T., Nusillard, B., Parmain, G., Soldati, F. & Bouget, C. (2015) PASSIFOR: A reference library of DNA barcodes for French saproxylic beetles (Insecta, Coleoptera). Biodiversity Data Journal, 3: e4078. doi: 10.3897/BDJ.3.e4078.
Smith, M.A., Bertrand, C., Crosby, K., Eveleigh, E.S., Fernandez-Triana, J., Fisher, B.L., Gibbs, J., Hajibabaei, M., Hallwachs, W., Hind, K., Hrcek, J., Huang, D.-W., Janda, M., Janzen, D.H., Li, Y., Miller, S.E., Packer, L., Quicke, D., Ratnasingham, S., Rodriguez, J., Rougerie, R., Shaw, M.R., Sheffield, C., Stahlhut, J.K., Steinke, D., Whitfield, J., Wood, M., & Zhou, X. (2012) Wolbachia and DNA Barcoding Insects: Patterns, Potential, and Problems. PLoS ONE, 7 (5): e36514. doi:10.1371/journal.pone.0036514
Takizawa, H. & Nakane, T. (1977) Byrrhus Linné and Cytilus Erichson (Coleoptera, Byrrhidae) from Japan and its Adjacent Areas. Bulletin of the National Museum of Nature and Science Series A (Zoology), 3 (1): 55-65.
Tshernyshev, S.E. (2006) A review of pill beetles (Coleoptera: Byrrhidae) of the fauna of Russia and the adjacent territories. Taxonomic composition. Proceedings of the Russian Entomological Society. St. Petersburg, 77: 287-293.
Tronquet, M. & Peslier, S. (2024) Catalogue des Coléoptères de France. Édition actualisée en temps réel [mars 2024]. Revue Roussillonnaise d’Entomologie, Perpignan, 867 pp.
Vuataz, L., Sanchez, A., Wyler, S., Blanc, M., & Chittaro, Y. (2019) Diversity and relationships of Ampedini Gistel, 1848 (Coleoptera: Elateridae) in Switzerland and Europe. Invertebrate systematics, 33 (3): 544-555.
B. Dodelin, 18 octobre 2024
(mise à jour de forme 10 janvier 2025)

Une réflexion au sujet de « Il y a bien deux espèces de Cytilus en Europe »